- Category: Volume 6
- Hits: 10898
Variation annuelle des paramètres éco-physiologiques de Argania spinosa L. Skeels var. mutica et var. apiculata dans le semi-aride tunisien
| [Volume 6, Article 1] | 496 kB |
S. El Adib ¹*
S. slim 2
F. Ben Jeddi ¹
1 Université de Carthage. Laboratoire des Sciences Horticoles, INA Tunisie,
2 Université de Carthage. Laboratoire des Sciences Horticoles, ESA Mateur.
Abstract - The Argan tree (Argania spinosa L. Skeels) contains two varieties, mutica (spineless) and apiculata (thorny). The seasonal effect on the chlorophylls concentrations, water relative contents (WRC), and leaf areas (LA) of argan trees in INAT arboretum were analyzed. During winter and spring seasons, a significant increase of the rates of leaf chlorophylls (a/b) was particularly found at A. apiculata. In january, the chlorophyll a concentrations were 2084.61 and 1605.05 µg/g MF (fresh material) respectively to apiculata and mutica. A. apiculata leaf area was reduced to 23.5 and 14.6 cm²/cm in october and january; compared to A. mutica which maintained a higher and relatively the same leaf area about to 35cm²/cm. In the same environment, leaves water relative content of A. apiculata reach a high level between 51.4 and 68.32% in february and march. Contrary to A. mutica, WRC has varied between 29.3 and 19.07%. During the dry season, A. apiculata is characterized by a photosynthetic activity more important than A. mutica with respectively 869.66 and 655.51µg/g MF of chlorophyll a.
Key words: Argania spinosa / water relative content / Chlorophylls / leaf area
Résumé - L’Arganier (Argania spinosa L. Skeels) comporte deux variétés, mutica (inerme) et apiculata (épineuse). L’effet saisonnier le long d’un cycle phénologique annuel sur la variation des concentrations en chlorophylles, teneurs relatives en eau, et surfaces foliaires de deux variétés d’arganier implantés dans l’arboretum de l’INA Tunisie a été analysé. Durant les saisons hivernale et printanière, une augmentation significative des taux de chlorophylles (a et b) dans les feuilles a été signalée particulièrement chez A. apiculata. Les concentrations en chlorophylle a du mois de janvier ont été de 2084,61 et 1605,05 µg/g MF (matière fraîche) respectivement chez A. apiculata et A. mutica. À la même période, la surface foliaire de A. apiculata a été réduite à 23,5 et 14,6 cm²/cm de rameau aux mois d’octobre et janvier. Contrairement à A. mutica qui a préservée une surface foliaire relativement stable et plus élevée de l’ordre de 35cm²/cm. Dans le même cadre spatio-temporel, la teneur relative en eau des feuilles de A. apiculata a marqué des niveaux élevés de 51,4 et 68,32 % en février et mars. Alors qu’elle a enregistré une chute significative chez A. mutica avec 29,3 et 19,07 %. Pendant la saison sèche, A. apiculata se caractérise par une activité photosynthétique plus importante que A. mutica avec respectivement 869,66 et 655,51 µg/g MF de chlorophylle a.
Mots clés : Argania spinosa / Teneur en eau relative / Chlorophylles / surface foliaire
1. Introduction
L’arganier (Argania spinosa L. Skeels) est une espèce endémique du sud ouest marocain (Emberger 1925; Boudy 1950; Ehrig 1974; M’hirit 1989; Prendergast et al. 1992). C’est un arbre thermophile et xérophile, avec des exigences en humidité atmosphérique élevées (Nouaim et Chaussod 1993). Les forêts d’arganier ne s’étendent actuellement que dans les zones arides et semi-arides du sud ouest marocain. En Tunisie, plusieurs essais d’introduction dans un grand nombre de stations ont été installés entre les années 1963 et 1968 (Hamrouni et al. 2009). La présence d’épines chez les sapotacées est un synonyme d’adaptation au stress hydrique. Les espèces des zones arides et semi-arides peuvent survivre au déficit hydrique et à l’extrême irrégularité des précipitations grâce à un ensemble de mécanismes morphologiques, anatomiques, physiologiques et métaboliques reflétant différents types d’adaptations (Turner 1986; Domingo et al. 2003; Sawadogo et al. 2006). Le passage du type inerme mutica à l’épineux apiculata confirme l’adaptation de l’arganier aux zones arides, phénomène confirmé par la diminution de la taille des feuilles. Lors d’un déficit hydrique, l’activité photosynthétique de la feuille se trouve affectée avec une diminution particulière de la concentration en chlorophylle (Kaya et al. 2006; Degl’Innocenti et al. 2008).
La surface foliaire correspond à une signification physiologique directe de l’état hydrique de l’arbre. Les espèces végétales à faible surface foliaire sont considérées comme tolérantes à la sécheresse. Dans cette optique, le travail s’intéresse à l’étude de certains paramètres physiologiques et de croissance de deux variétés d’arganier mutica et apiculata durant les quatre saisons de l’année dans les semi aride supérieur à hiver doux de la Tunisie.
2. Matériels et méthodes
2.1. Matériel biologique
Le matériel végétal est constitué de feuilles d’arganier prélevées de deux variétés d’arganier différentes inerme (mutica) et épineuse (apiculata) situées dans l’arboretum de l’INA Tunisie. L’arboretum de l’INAT appartient à l’étage bioclimatique semi-aride supérieur à hiver doux. Le sol est de type argilo-limoneux. Selon Google Earth (2011), les coordonnées géographiques des arbres étudiés sont respectivement 36°49’53.37’’N/10°10’52.9’’E et 36°49’55.5’’N/10°10’53.0’’E. Chaque arbre a été est divisé en quatre zones selon les orientations naturelles NE, NO, SE et SO afin d’homogénéiser les prélèvements des rameaux au nombre de trois par orientation et par mois répartis sur une année.
2.2. Extraction et dosage des pigments chlorophylliens a et b
Les teneurs des feuilles en chlorophylles a, b, et totale exprimées en µg/g MF (matière fraîche) ont été déterminées selon la méthode de Torrecillas (1984), 100 mg de feuilles fraîches et saines ont été pesées et mises dans 5 ml d’acétone à 80%. Après un séjour de 72 heures à l’obscurité et à une température de 4°C, la densité optique (DO) a été mesurée à 645 et 663 nm avec un spectrophotomètre UV-Visible (Labomed, Inc, USA). Les équations suivantes établies par Mc Kinney (1941) et Anon (1949), ont été utilisées pour calculer les teneurs en chlorophylles a, b et totale:
-
chlorophylle a: 12,7 x DO (663 nm) – 2,69 x DO (645 nm);
-
chlorophylle b: 22,9 x DO (645 nm) – 4,68 x DO (663 nm); et
-
chlorophylle totale: 20,2 x DO (645 nm) + 8,02 x DO (663 nm).
2.3. Surface foliaire
La surface foliaire est une mesure indicatrice de l’état du fonctionnement foliaire des arbres nécessaire à l’assimilation énergétique. Ce paramètre a été mesuré sur trois rameaux de 30 cm de longueur issus des quatre orientations par variété et par mois à l’aide d’un planimètre de marque LICOR. L’unité de mesure a été exprimée en cm²/cm de rameau.
2.4. Teneur relative en eau des feuilles
Les mêmes feuilles de la surface foliaire ont été emballées dans des paquets pour déterminer leur masse fraîche totale par une balance de précision. Ces paquets ont été placés par la suite dans une étuve à une température de 70°C pendant 72 heures. La masse sèche des feuilles a été déterminée avec une balance de précision.
TRE % = (poids frais-poids sec / poids frais) x 100
2.5. Analyses statistiques des données
Les différents paramètres étudiés ont été analysés moyennant le logiciel SAS (version 9.1). Ces analyses ont porté sur la variance en utilisant la procédure PROC ANOVA. La comparaison des moyennes a été effectuée selon la méthode LSD au seuil 5%. La signification de la variance a été vérifiée par le test de Fisher.
3. Résultats
3.1. La chlorophylle a de l’arganier
Le dosage mensuel de la chlorophylle a dans les feuilles d’arganier a montré une divergence significative entre les mois et particulièrement les saisons. Pour les deux variétés apiculata et mutica, l’augmentation de la chlorophylle a (figure1) a commencée au mois de novembre avec des concentrations similaires pour les deux variétés 1220 et 1261 µg/g MF pour atteindre son maximum au mois de mars avec 2101,41 et 1459,14 µg/g MF respectivement pour apiculata et mutica (figure 1). Une diminution significative a été notée à partir du mois d’avril et le long de la saison estivale. Les feuilles de la variété apiculata ont présenté des teneurs assez importantes que la variété mutica en matière de chlorophylle a au cours des mois et notamment à partir de décembre.
|
|
|
Figure 1: Variation annuelle de la chlorophylle a dans les feuilles des deux variétés d’arganier mutica (a) et apiculata (b) |


3.2. La chlorophylle b de l’arganier
Le dosage de la chlorophylle b dans les feuilles d’arganier le long du même cycle phénologique a illustré une dynamique saisonnière relativement similaire à celle de la chlorophylle a (figure 2). Des différences significatives entre les mois ont été notées. Ainsi, l’augmentation des teneurs en pigments chlorophylliens a commencée au mois de décembre avec des différences non significatives entre les deux variétés. Un premier pic est apparu au mois de Novembre avec 1261,07 µg /g MF. Le maximum de synthèse a été obtenu en février avec 1659,32µg/g MF. La variété apiculata est la plus dynamique en mode de synthèse chlorophyllienne surtout en hiver et au printemps par rapport à la variété mutica.
|
|
|
Figure 2 : variation annuelle de la quantité de chlorophylle « b » dans les feuilles des deux variétés mutica (a) et apiculata (b) le long d’un cycle phénologique |


3.3. La chlorophylle totale le long d’un cycle phrénologique
L’augmentation de la synthèse des pigments a commencée au mois d’octobre avec des teneurs similaires entre les feuilles des deux variétés apiculata et mutica (figure 3).Une première fluctuation au mois de décembre et une deuxième au mois de février avec des différences significatives dans la réponse des deux variétés. Une baisse spectaculaire au mois de mars surtout pour la variété épineuse suivie d’une augmentation au mois d’avril. Les taux restent faibles pour les deux variétés pendant la saison estivale mais ils sont plus importants dans les feuilles de la variété apiculata.
|
|
|
Figure 3 : variation annuelle de la quantité de chlorophylle totale dans les feuilles des deux variétés mutica (a) et apiculata (b) le long d’un cycle phénologique |
3. 4. Etude de la dynamique de la teneur relative en eau des feuilles de l’arganier
La teneur relative en eau des feuilles de la variété apiculata est élevée le long du cycle phénologique (entre 45 et 60%). Les variations sont négligeables entre les mois et les différences sont non significatives (figure 4). Pour la variété mutica, la dynamique de la variation de la pression osmotique des feuilles d’une saison à l’autre marque des différences significatives. Ainsi les teneurs sont relativement élevées en l’automne avec 58 et 57% respectivement en octobre et novembre. Alors que des baisses remarquables ont commencées en décembre et janvier (48 et 41%) pour continuer jusqu’à la fin du printemps avec 16% en mai. Une augmentation commence à apparaitre en août avec une teneur relative en eau de 47% qui représente presque le double de celle de mai, juin et Juillet.
|
|
|
Figure 4: Variation du pourcentage de la teneur relative en eau TRE dans les feuilles des deux variétés mutica (a) et apiculata (b) le long d’un cycle phénologique de 12 mois |


3 .5. Etude de la dynamique de la variation de la surface foliaire des feuilles des deux variétés
La surface foliaire des feuilles de la variété mutica est élevée le long du cycle phénologique entre 23 et 37 cm²/cm de rameau. Les variations sont d’ordre minime entre les mois et les différences sont non significatives entre les saisons (figure 5). Pour la variété apiculata, on marque une fluctuation dans la taille des feuilles d’une saison l’autre avec des différences significatives entre les saisons. Les indices foliaires sont élevés en automne avec 23,5 et 21,95 cm²/cm respectivement en octobre et novembre. Des baisses remarquables ont commencée en décembre et janvier avec 18,4 et 14,6 cm²/cm de rameau jusqu’à la fin du printemps avec 7,8 cm²/cm en mai et avril. Une augmentation commence à apparaitre en Aout avec un indice foliaire de 18,4 cm²/cm de rameau qui représente le double de la surface foliaire des feuilles moyenne des mois de mai, juin et juillet.
|
|
|
Figure 5 : variation de la surface foliaire (en cm²/cm de rameau) des feuilles des deux variétés mutica (a) et apiculata (b) le long d’un cycle phénologique de 12 mois |


|
Tableau 1-a : Résultats d’analyses statistiques des taux de chlorophylles dans les feuilles d’arganier variété mutica en µg/g MF |
|||
|
|
chlorophylle a |
chlorophylle b |
chlorophylle Totale |
|
|
|
|
|
|
septembre |
1032,3± 438,3 dc |
322,9±62,3cd |
1274,4±219,4 de |
|
|
|
|
|
|
Octobre |
828,8±197,4 de |
385,6±87,1 bc |
1287,2±228,5de |
|
|
|
|
|
|
Novembre |
1261,1±86,3 bc |
669,2±74,5 a |
2013,5±231,9bac |
|
|
|
|
|
|
Décembre |
1428,8±102,8 ba |
609,2±18,1 a |
2065,6±70,2 ba |
|
|
|
|
|
|
Janvier |
1605,1±159,a |
650,5±72,8 a |
2310,6±229,3 a |
|
|
|
|
|
|
Février |
1659,3±150,8 a |
693,2±48,6 a |
2403,4±201,5 a |
|
|
|
|
|
|
Mars |
1459,1±136,3 ba |
541,7±136,2 ba |
1948,4±387,1bac |
|
|
|
|
|
|
Avril |
1506,5±160,6ba |
525,4±43,6 ba |
1727,4±141,6bdc |
|
|
|
|
|
|
Mai |
925,8±87,42 de |
207,0±72dc |
982,7±162,6 e |
|
|
|
|
|
|
Juin |
655,5±154,8 e |
132,8±32 d |
810,1±161 e |
|
|
|
|
|
|
Juillet |
943,1±162,7 de |
547,6±54,3 ba |
1294,4±254,7 ed |
|
|
|
|
|
|
Aout |
1075,8±203,2cd |
543,9±79,6 ba |
1518,7±234,5 dc |
|
Les moyennes dans chaque colonne suivies par une lettre différente sont significativement différentes (p < 0,05). |
|||
|
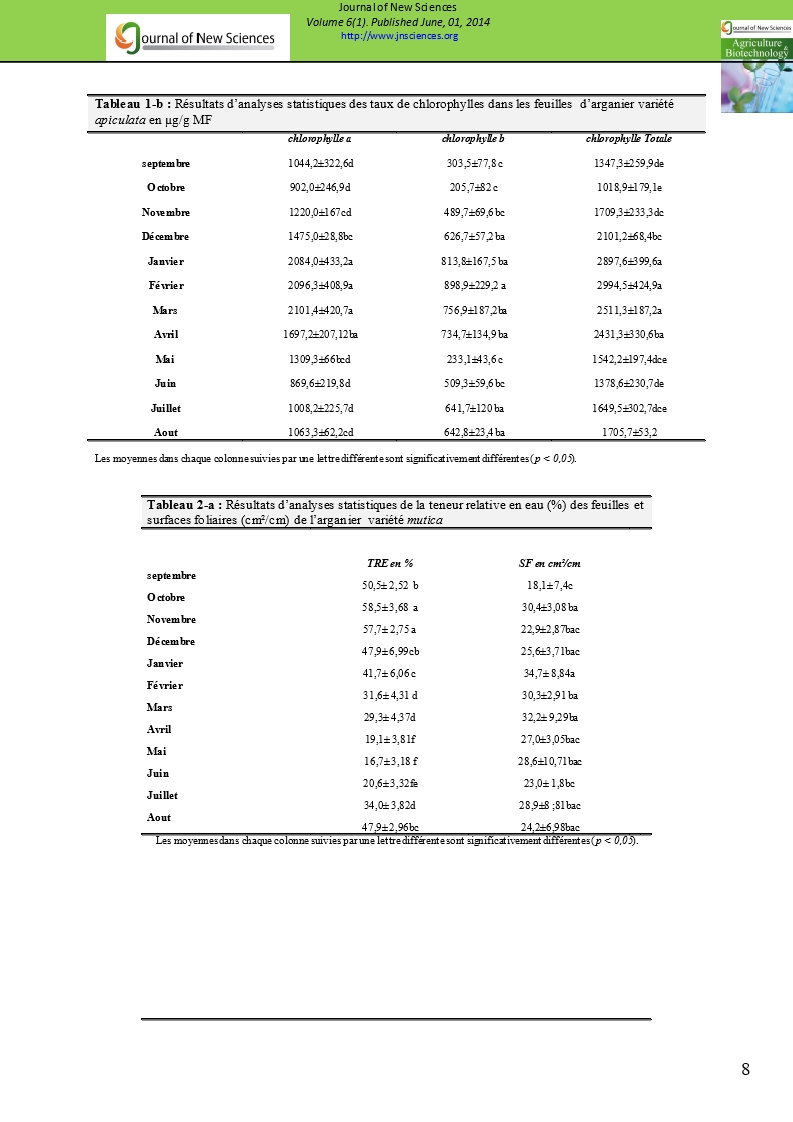
Tableau 1-b : Résultats d’analyses statistiques des taux de chlorophylles dans les feuilles d’arganier variété apiculata en µg/g MF |
|||
|
|
chlorophylle a |
chlorophylle b |
chlorophylle Totale |
|
|
|
|
|
|
septembre |
1044,2±322,6d |
303,5±77,8 c |
1347,3±259,9de |
|
|
|
|
|
|
Octobre |
902,0±246,9d |
205,7±82 c |
1018,9±179,1e |
|
|
|
|
|
|
Novembre |
1220,0±167cd |
489,7±69,6 bc |
1709,3±233,3dc |
|
|
|
|
|
|
Décembre |
1475,0±28,8bc |
626,7±57,2 ba |
2101,2±68,4bc |
|
|
|
|
|
|
Janvier |
2084,0±433,2a |
813,8±167,5 ba |
2897,6±399,6a |
|
|
|
|
|
|
Février |
2096,3±408,9a |
898,9±229,2 a |
2994,5±424,9a |
|
|
|
|
|
|
Mars |
2101,4±420,7a |
756,9±187,2ba |
2511,3±187,2a |
|
|
|
|
|
|
Avril |
1697,2±207,12ba |
734,7±134,9 ba |
2431,3±330,6ba |
|
|
|
|
|
|
Mai |
1309,3±66bcd |
233,1±43,6 c |
1542,2±197,4dce |
|
|
|
|
|
|
Juin |
869,6±219,8d |
509,3±59,6 bc |
1378,6±230,7de |
|
|
|
|
|
|
Juillet |
1008,2±225,7d |
641,7±120 ba |
1649,5±302,7dce |
|
|
|
|
|
|
Aout |
1063,3±62,2cd |
642,8±23,4 ba |
1705,7±53,2 |
|
Les moyennes dans chaque colonne suivies par une lettre différente sont significativement différentes (p < 0,05). |
|||
|
Tableau 2-a : Résultats d’analyses statistiques de la teneur relative en eau (%) des feuilles et surfaces foliaires (cm²/cm) de l’arganier variété mutica |
||||||||
|
|
|
|
||||||
|
TRE en % |
SF en cm²/cm |
|
|
|||||
|
septembre |
50,5± 2,52 b |
18,1± 7,4c |
|
|
||||
|
Octobre |
58,5± 3,68 a |
30,4±3,08 ba |
|
|
||||
|
Novembre |
57,7± 2,75 a |
22,9±2,87bac |
|
|
||||
|
Décembre |
47,9± 6,99cb |
25,6±3,71bac |
|
|
||||
|
Janvier |
41,7± 6,06 c |
34,7± 8,84a |
|
|
||||
|
Février |
31,6± 4,31 d |
30,3±2,91 ba |
|
|
||||
|
Mars |
29,3± 4,37d |
32,2± 9,29ba |
|
|
||||
|
Avril |
19,1± 3,81f |
27,0±3,05bac |
|
|
||||
|
Mai |
16,7± 3,18 f |
28,6±10,71bac |
|
|
||||
|
Juin |
20,6± 3,32fe |
23,0± 1,8bc |
|
|
||||
|
Juillet |
34,0± 3,82d |
28,9±8 ;81bac |
|
|
||||
|
Aout |
47,9± 2,96bc |
24,2±6,98bac |
|
|
||||
|
Les moyennes dans chaque colonne suivies par une lettre différente sont significativement différentes (p < 0,05).
|
||||||||
|
Tableau 2-b : Résultats d’analyses statistiques de la teneur relative en eau (%) des feuilles et surfaces foliaires (cm²/cm) de l’arganier variété apiculata |
||||||||
|
|
|
|
||||||
|
TRE en % |
SF en cm²/cm |
|
|
|||||
|
septembre |
58,0± 5,51a |
23,1 ±2,15a |
|
|
||||
|
Octobre |
57,2± 1,83a |
23,5±4,45 a |
|
|
||||
|
Novembre |
54,14±7,56 a |
20,2± 1,27ba |
|
|
||||
|
Décembre |
58,8±16,71 a |
18,4±2,91 bc |
|
|
||||
|
Janvier |
32,7± 1,75a |
14,6± 2,77cd |
|
|
||||
|
Février |
48,5± 7,0a |
12,8± 3,19ed |
|
|
||||
|
Mars |
51,4±29,06 a |
9,8±2,32 fe |
|
|
||||
|
Avril |
68,3± 6,96a |
7,8± 1,64f |
|
|
||||
|
Mai |
58,2±4,41 a |
7,7± 2,27f |
|
|
||||
|
Juin |
60,3±1,67 a |
12,8± 0,77ed |
|
|
||||
|
Juillet |
56,1± 1,84a |
16± 4,38bcd |
|
|
||||
|
Aout |
57,6± 1,68a |
19,3±2,38 ba |
|
|
||||
|
Les moyennes dans chaque colonne suivies par une lettre différente sont significativement différentes (p < 0,05). |
||||||||
4. Discussion
Chez les deux variétés d’arganier mutica et apiculata, la dynamique de la synthèse chlorophyllienne le long du cycle phénologique est généralement proportionnelle au cours des saisons mais avec des différences significatives en quantité de chlorophylles (chl). La teneur des pigments chlorophylliens (chl a, chl b et chl totale) diminue selon un gradient d’aridité croissant (Bouzoubaa et al. 2011). La synthèse de la chlorophylle baisse durant les saisons sèches suite aux contraintes hydriques. D’après Lowlor (2002), l’activité physiologique de la feuille et plus particulièrement la photosynthèse et la conductance stomatique sont affectées lors d’un déficit hydrique (Lawlor et Cornic 2002). Chez les deux variétés, avec l’augmentation de la température, la synthèse de la chlorophylle b se trouve affectée en premier lieu et par la suite la chlorophylle a. La réduction de la photosynthèse, liée à la diminution du potentiel hydrique foliaire, dépend à la fois de la fermeture des stomates, avec une diminution de la conductance à la diffusion du CO2 et d’une limitation biochimique du chloroplaste à le fixer (Tardieu et Simoneau 1998; Escolana et al. 1999; Flexas et Medrano 2002). En saison sèche, la synthèse de la chlorophylle dans les feuilles de A. apiculata est plus significative que celle de A. mutica. Cette variété épineuse serait mieux adaptée au stress hydrique suite à la réduction de sa surface foliaire (Parry et al. 2005; Lebon et al. 2006) pour pouvoir limiter l’évapotranspiration. Cette réduction de la surface foliaire peut provenir d’une diminution de l’expansion foliaire et/ou d’une sénescence accélérée de la feuille (Kramer et Boyer 1995).
La croissance foliaire est stoppée très rapidement par un déficit hydrique ce qui explique la taille réduite des feuilles de A. apiculata. Cette stratégie est principalement liée, d’une part, à la réduction de la transpiration et d’autre part, à une optimisation de l’absorption d’eau par les racines. La diminution de la transpiration est principalement liée à la fermeture des stomates. Elle peut être due également à une diminution des surfaces évaporantes. La régulation de la conductance stomatique reste le mécanisme majeur intervenant à court terme pour limiter les pertes en eau. Le potentiel hydrique foliaire sera maintenu d’autant plus longtemps que la fermeture des stomates est précoce. La régulation de la conductance stomatique chez les sapotacées dépend du potentiel hydrique foliaire et de l’humidité de l’air au champ (Lawlor et Cornic 2002). Une forte corrélation entre la dynamique de synthèse de la chlorophylle d’une part et TRE et IF pour chaque variété d’une autre part a été observée. Chez A. apiculata, le maintien de taux relativement élevé en matière de chlorophylle par rapport à A. mutica en condition de contrainte hydrique n’est que la conséquence d’une baisse de sa surface foliaire. D’autre part, A. mutica n’a pas réagit à une insuffisance hydrique tout en maintenant une surface foliaire relativement élevée le long du cycle. L’activité photosynthétique est totalement influencée par la teneur relative en eau des feuilles ainsi chez la variété épineuse, la synthèse de chlorophylle reste toujours meilleure que celle de la mutica grâce au maintient des réserves en eau dans les feuilles Un comportement qui a affecté directement les réserves hydriques des feuilles en provoquant une perturbation photosynthétique et donc une diminution de la synthèse chlorophyllienne (Sabater et al. 1978). La variété apiculata subit une contrainte hydrique alors que mutica subit un déficit hydrique. Selon (Lecoeur et al. 2007), si le fonctionnement de la plante est affecté par un déficit hydrique mais les tissus de la plante ne subissent pas une baisse du TRE, on parlera alors de contrainte hydrique, mais on parle de stress hydrique lorsque les tissus de la plante subissent une baisse de leur teneur relative en eau qui affecte le métabolisme de la plante et spécifiquement la photosynthèse qui est le cas de la variété mutica. Une forte corrélation entre la TRE et la surface foliaire pour les deux variétés ainsi, pour toute diminution de la surface foliaire on remarque une bonne rétention d’eau dans les feuilles et cela est tout a fait logique car la diminution de la surface foliaire induit la fermeture des stomates et donc la limitation de l’évapotranspiration (EL Aboudi 1990). Le déficit hydrique pourra être le résultat d’une faible disponibilité en eau dans le sol ou une faible exploitation des réserves hydriques disponible dans le sol par les racines et/ou d’une forte demande évaporatoire au niveau des feuilles c'est-à-dire que l’évapotranspiration maximale ETM est plus élevée que la transpiration (Gadallah 1995). La variété apiculata est plus adaptée aux conditions de stress hydrique ainsi elle pratique la stratégie d’évitement pour minimiser la transpiration et la métamorphose des feuilles en épines aussi pour minimiser l’évapotranspiration. De ce faite, tout les processus de la plante seront affectés et principalement le processus métabolique par la réduction de la photosynthèse et le processus de l’organogenèse par la réduction de la taille des fruits et de leurs propriétés phytochimiques (Thapar et al. 2008).
5. Conclusion
L’ensemble des résultats montre que les deux variétés d’arganier sont généralement tolérantes à la sécheresse. La variété apiculata s’est montrée plus adaptée aux conditions de stress hydrique grâce à la stratégie d’évitement en limitant la transpiration par la fermeture partielle des stomates d’une part et la rétention du maximum d’eau dans ses feuilles d’autre part. Malgré une surface foliaire plus grande par rapport à la variété apiculata, la variété mutica est moins active en matière de photosynthèse surtout avec l’augmentation de la température. Pour les deux variétés, la synthèse des pigments chlorophylliens dépend totalement de l’état physique et hydrique des feuilles indépendamment de la saison.
6. Références
Arnon DI (1949) Copper Enzymes in isolated chloroplasts, polyphenoxidase in Beta Vulgaris. Plant physiology 24. pp: 1-15.
Bota J, Flexas J et Medrano H (2001) Genetic variability of photosynthesis and water use in Balearic grapevine cultivars. Annals of Applied Biology 138. pp: 353-361.
Baker NR and Rosenqvist E (2004) Application of chlorophyll fluorescence can improve crop production strategies an examination of future possibilities. Journal of Experimental Botany 55. pp: 1607-1621.
Boudy P (1950) Economie forestière Nord Africaine. Tome II, Monographie et traitements des essences forestières. Fasc 1. pp: 382-416. Larose. Paris.
Domingo F, Brenner AJ, Gutiérrez L, Clark SC, Incoll LD et Aguilera C (2003) Water relations only partly explain the distributions of three perennial plant species in a semi-arid environment. Biologia plantarum 46. pp: 257-262.
Degl’Innocenti E, Guidi L, Stevanovic B, Navari F (2008) CO2 fixation and chlorophyll a fluorescence in leaves of Ramonda serbica during a dehydration–rehydration cycle. Journal of Plant Physiology 165. pp: 723-733.
Demming-Adams B and Adams W (1996) The role of xanthophyll cycle carotenoides in the protection of photosynthesis. Tends plant Sci I. pp: 21-26.
Ehrig FR, (1974) Die Arganie. Charakter, Ökologie und Witschaftliche Bedeutung eines Teträrreliktes in Marokko. Petermanns Geogr. Mitt 118. pp: 117-125.
El Aboudi A, (1990) Typologie des agraneraies inframediterraneennes et ecophysiologie de l’arganier (Argania spinosa (L.) Skeels) dans le Sous (Maroc). Thèse de doctorat. Université JosephFourier Grenoble I: p123.
Escalona JM, Flexas J et Medrano H, (1999) Stomatal and non-stomatal limitations of photosynthesis under water stress in field-grown grapevines. Australian Journal of Plant Physiology 26.pp: 421-433.
Emberger L, (1925) Le domaine naturel de l’arganier. Bull. Soc. Bot., Paris. pp: 770-774.
Flexas J et Medrano H, (2002) Drought-inhibition of photosynthesis in C3 plant: Stomatal and non-stomatal limitations revisited. Annals of Botany 89. pp: 183-189.
Gadallah M A A, (1995) Effect of water stress, abscisic acid and proline on cotton plants. Journal of Arid Environment 30. pp: 315-325.
Gimenez C, Mitchell VJ et Lawlor DW, (1992) Regulation of photosynthetic rate of two sunflower hybrids under water stress. Plant Physiol 98.pp: 516-524.
Guan XQ, Zhao SJ, Li DQ and Shu HR, (2004) Photoprotective function of photorespiration in several grapevine cultivars under drought stress. Photosynthetica 42. pp: 31-36.
Howell GS, (2001) Sustainable grape productivity and the growth-yield relationship: A review. Amer J. Enol. Vitic. 52. pp: 165-174.
Kaya C, Tuna AL, et Alves AAC, (2006) Gibberellic acid improves water deficit tolerance in maize plants. Acta physiologiae plantarum. 28. pp: 331-337.
Kramer PJ et Boyer JS, (1995) Water relations of plants and soils (Book). Academic Press, Inc 19. pp: 123-132.
Lawlor DW et Cornic G, (2002) Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell & Environment 25. pp: 275-294.
Lebon E, Pellegrino A, Tardieu F et Lecoeur J, (2004) Shoot development in grapevine is affected by the modular branching pattern of the stem and intra and inter-shoot trophic competition. Annals of Botany 93.pp: 263-274.
Lebon E, Pellegrino A, Louarn G et J Lecoeur J, (2006) Branch development controls leaf areadynamics in grapevine (Vitis vinifera) growing in drying soil. Annals of Botany 98. pp: 175-185.
Lecoeur J, Wery J, Turc O et Tardieu F, (1995) Expansion of pea leaves subjected to short water deficit: cell number and cell size are sensitive to stress at different periods of leaf development. Journal of Experimental Botany 46. pp: 1093-1101.
Lecoeur et al (2007) Influence d’un déficit hydrique sur le fonctionnement d’un couvert végétal cultivé. INRIA research Report.
Le Maire R, (1929) Contributions à l’étude de la flore de l’afrique du nord, (fascicule 14). Bull. Soc. Hist. Nat. Afr. Nord, 20. pp: 12 à 42.
Mackinney G, (1941) Absorption of light of chlorophyll solution. J. bio.Chem 140. pp: 315-322.
M’hirit O, (1989) L’arganier est une espèce fruitière forestière à usages multiples: Formation forestière continue, Thème « l’arganier » Station de Recherche Forestière, Rabat, 13-17 Mars. pp: 31-57.
Nouaim R, Chaussod R, (1993) L’arganier – Argania spinosa (L) Skeels.(Sapotaceées). Bull Res Arb Trop 27 .pp: 7-9.
Parry MAJ, Flexas J et Medrano H, (2005) Prospects for crop production under drought: research priorities and future directions. Annals of Applied Biology 147. pp: 211-226.
Prendergast HDV, Walker CC, (1992) The Argan multipurpose tree of Morocco. The Kew Magazine 9. pp: 76–85.
Sabater B , Rodriquez MI, (1978) Control of chlorophyll degradation in detached leaves of barley and oat through effect of kinetin on chlorophyllase levels. Physiol Plant 43. pp: 274-276
Sawadogo M, Zombre G, Balma D, (2006) Expression de différents écotypes de gombo (Abelmoschus esculentus L.) au déficit hydrique intervenant pendant la boutonnisation et la floraison.Biotechnol Agron Soc Environ. pp: 43-54.
Soar CJ, Speirs J, Maffei SM, Penrose AB., McCarthy MG. and Loveys BR, (2006) Grape vine varieties Shiraz and Grenache differ in their stomatal response to VPD: apparent links with ABA physiology and gene expression in leaf tissue. Australian Journal of Grape & Wine Research 12. pp: 2-11.
Tardieu F, Zhang J and Gowing DJG, (1993) A model of stomatal control by both ABA concentration in the xylem sap and leaf water status:Test of the model and of alternative mechanisms for droughted and ABA-fed field-grown maize. Plant Cell & Environment 16. pp: 413-420.
Tardieu F, (2005) Plant tolerance to water deficit: physical limits and possibilities for progress. C. R. Geoscience 337. pp: 57-67.
Thapar, R, Srivastava AK, Bhargava P, Mishra Y et Rai LC, ( 2008) Impact of different abiotic stress on growth, photosynthetic electron transport chain, nutrient uptake and enzyme activities of Cu-acclimated Anabaena doliolum. J Plant Physiol 165. pp: 306-316.
Tardieu F and Simoneau, (1998) Variability among species of stomatal control under fluctuating soil water status and evaporative demand: modeling isohydric and anisohydric behaviours. Journal of Experimental Botany 49. pp: 419-432.
Turner NC, (1986) Adaptation to water deficits: A changing perspective. Aust J Plant Physiol 13. pp: 175-90.
