- Category: Volume 7
- Hits: 13746
Watering regime affects oil content and fatty acid composition of six sunflower lines
| [Volume 7, Article 1] | 170 kB |
Sahari Khoufi1*
Khalil Khamassi1
Jaime A. Teixeira da Silva2
Salah Rezgui1
Fayçal Ben Jeddi1
1 Unité Cultures Maraîchères et Florales, Institut National Agronomique de Tunisie, 43, avenue Charles Nicolle 1082 Tunis-Mahrajène Tunisie. Université de Carthage, Tunisia
2 P.O. Box 7, Miki cho post office, Ikenobe 3011-2, Kagawa-Ken, 761-0799, Japan.
Abstract - Six sunflower lines were grown under field conditions in order to assess their oil content and fatty acid composition under three watering regimes (rain-fed, 50 % supply, and 100 % supply). Changes in oil percentage and fatty acid composition of sunflower seeds were determined and analyzed using a Soxhlet apparatus and gas chromatography. A significant genotype × water regime interaction for oil content was noted. Contents of linoleic acid and stearic acid were affected only by watering regime and ranged from 4.86 to 12.33 % for the former and from 1.90 to 3.21 % for the latter. Palmitic oil content was affected by genotype: 6.03 %, 7.54 %, 4.18 %, 4.40 %, 5.05 % and 4.52 % for C8.3.1.2, 64A71, C7.3.2.2, C1.3.1.1, C5.2.3.2, and C31.1.1.2, respectively. Oleic content was independent of the effects imposed by genotype and water regime.
Key words: oil content / saturated fatty acids / sunflowers / unsaturated fatty acids / water supply / Soxhlet.
1. Introduction
Sunflower (Helianthus annuus L.) is an important oilseed crop whose oil content varies from 25 to 50 % of seed content (Izquierdo et al. 2002). Sunflower oil contains large amounts of unsaturated fatty acids, mainly linoleic (C18:2) and oleic acid (C18:1) (14 to 38 % and 46 to 74 %, respectively). Lower levels of saturated fatty acids in sunflower are mainly represented by palmitic acid (C16:0) and stearic acid (4.4 to 7.7 % and 2.8 to 7.5 %, respectively) (Harwood and Aparicio 2000). Different studies (Table 1) have shown that the composition of fatty acids and oil seed content depends on genotype. Based on the composition of fatty acids, CODEX Stan 210 (1999) classifies sunflower into three categories. The production of linoleic acid ranged from 48 to 74 % in standard sunflower in comparison to greater oleic acid production (75-85 %) noted in high oleic hybrid genotypes and 50-75 % in oleic genotypes (Anastasi et al. 2000). The Institut Français des Corps Gras (ITERG) showed that the French variety ‘Mirasol’ contains 19.8 % of oleic acid and 67.0 % of linoleic acid (Boyeldieu 1991). Pectu et al. (2001) reported that Romanian sunflower had more than 60 % and 28-35 % linoleic acid and oleic acid, respectively.
Variation in oil content and composition has been attributed to climatic conditions prevailing during the grain-filling period and maturity. For example, Anastasi et al. (2000) and Piva et al. (2000) reported that temperature had no significant effect on oleic and linoleic acid. Conversely, several studies reported that the ratio of oleic/linoleic acid increases under high temperature during seed maturation and decreases under lower temperature (Anastasi et al. 2000; Rondanini et al. 2003; Roche et al. 2006; Rondanini et al. 2006; Pereira-Irujo and Aguirrezábal, 2007; Izquierdo and Aguirrezábal, 2008; Echarte et al. 2010). Baldini et al. (2002) and Flagella et al. (2004) demonstrated that both saline and drought stresses decreased seed oil content in hybrids with high oleic acid content. However, the magnitude of this decrease was associated with the intensity of the stress and the time when the stress occurred. The relation between fatty acid composition and drought stress in sunflower remains poorly explained and studies on this topic are contradictory. Baldini et al. (2002) and Flagella et al. (2000) found that water stress increased oleic acid content and decreased linoleic acid content in both standard and high oleic genotypes while Flagella et al. (2002), Santonoceto et al. (2003) and Pectu et al. (2001) reported that water stress decreased oleic acid content and increased linoleic acid content in standard sunflower hybrids by as much as 14 %. Saturated fatty acids like palmitic acid (C18:0) may increase in response to water stress (Piva et al. 2000) while Santonoceto et al. (2003) and Pectu et al. (2001) noted no significant change in saturated fatty acids, mainly palmitic and stearic acids.
The objectives of this study were to determine the changes in seed oil content and fatty acid composition, i.e., of saturated and unsaturated lipids, of six sunflower genotypes cultivated in Tunisia exposed to three watering regimes.
2. Materials and methods
Six adapted sunflower lines (Helianthus annuus L.) were grown in an experimental field at the Agronomic National Institute of Tunisia in 2011. These lines were provided by the National Agronomic Institute of Tunisia. In the experimental field, seeds were planted, in March, in a randomized block design with three replications. Seeds were planted manually, each block included 6 rows to accommodate the 6 lines, and three water regimes were applied: 1) 125 mm (natural rainfall = rainfed); 2) 50 % of water needs (RH50 = 310 mm); and 3) optimal needs (100 %) (RH100 = 510 mm). Regimes 2 and 3 were based on prior recommendations for sunflower (Blanchet et al. 1990; Centre Technique Interprofessionnel des 95 Oléagineux Métropolitains and La Filière Française des Huiles Et Protéines Végétales, 2008).
Analysis of the total oil content was performed by n-hexane extraction using a Soxhlet apparatus (Association Française de Normalisation, 1998). Mature achenes (20 g) were ground in an electric grinder and introduced into cartridges made of thick filter paper. Oil content was extracted separately from ground achenes of each genotype with n-hexane over 8 h. Then, the solvent was removed from the extracts under low pressure evaporation using a rotary evaporator (Büchi-Rotavapor RIIIHP, Büchi Labortechnik, Switzerland) and lipid extracts were weighed.
|
Table 1: Studies related to the effect of water supply on oil content, and saturated and unsaturated fatty acids of sunflower (Helianthus annuus L.). |
|||
|
Reference |
Genotypes and country of study |
Stress conditions |
Stress effect on oil content and fatty acids (saturated and unsaturated acids) |
|
Salera and Baldini 1998 |
Hybrids of sunflowers (high oleic and low oleic genotypes). Mediterranean climate. |
Different environmental conditions. |
No effect on oleic acid content and OA/LA ratio on high oleic hybrids. Low effect on low oleic hybrids. |
|
Piva et al. 2000 |
Sunflower genotypes. Mediterranean climate. |
Water deficit during flowering to seed filling stage. |
Saturated fatty acids increase (P and S). |
|
Baldini et al. 2002 |
Standard and oleic genotypes. Italy. |
Discontinuous irrigation regimes during the crop cycle. Flowering, seed filling (early stress). Seed-filling, maturity (late stress) |
Oil content decreased (9 %). OA/LA ratio decreased (2.09 %) in early stress. OA/LA ratio decreased (1.52 %) in late stress. |
|
Flagella et al. 2002 |
High oleic hybrids (Platon and Vyp70). Italy. |
Irrigation regime in critical stages (bud flowering appearance and flowering). Non-irrigated regime (rainfall). |
Rainfall regime increases SA and oil yield. Irrigation regime decreases SA and OA content (2.6 %), increases PA and LA (2.8 %). |
|
Santonoceto et al. 2003 |
Sunflowers genotypes. Italy. |
Water regimes: (33 % to 100 % of water needs of sunflower). |
Oil content increases with increasing water supply. No effect on saturated fatty acids. |
|
Roche 2005 |
Oleic genotypes. France. |
Regime 70% of water needs of sunflower. Rainfall regime. |
Oil content decreases in severe stress conditions. SA, PA and LA increase in severe stress conditions. |
|
Maksimović 2005 |
Sunflowers hybrids (Dukat and NS-H1-11). Serbia. |
Irrigation regime was scheduled on the basis of soil moisture, at 60-65% of field water capacity. |
Oil content increases. |
|
Kazemeini et al., 2009 |
Sunflower seeds (var. Euroflor). Iran. |
I2 and I3: 75 and 50 % soil water content in field capacity (FC), (Stem elongation maturity). I4 and I5: 75 and 50 % FC, (Flowering maturity). Control (I1: full irrigation and increasing soil moisture content in rhizosphere to field capacity). |
Reduced irrigation water during flowering decrease oil yield. |
|
Anastasi et al., 2010 |
Standard genotypes (Gs) and oleic genotype (Go). Italy. |
The water supply consist of four level: 0% (ETc0), 33 % (ETc33), 67 % (ETc67) and 100 % (ETc100) throughout the entire crop cycle. |
In Gs PA decreases, SA increases and OA/LA ratio decreases in increasing water stress condition. In Go PA and SA are stable and OA/LA ratio increases in increasing water stress condition. |
|
Alahdadi et al., 2011 |
Hybrids: Azargol, Alstar, Hysun 33 and Hysun 25. Iran |
Three irrigation regimes consisting of normal irrigation, mild stress and intense stress corresponding to 50, 100 and 150 mm cumulative evaporation respectively. |
Oil content decreases in all hybrids. The decrease depends on genetic base of each hybrid. SA increases under drought conditions. |
|
Sazen et al., 2011 |
Pioneer-64A14. Turkey |
Five irrigations consisting of: I1 = full irrigation. I2, I3 and I4, deficit irrigation. I5 = rainfall regime. |
Oil content increase with increasing irrigation. Increasing irrigation increases LA and decreases OA. |
|
Kaya and Kolsarici 2011 |
Hybrids (Sanbro, Tarsan-1018 and Ozdemirbey). Turkey. |
Seven irrigation schedules; I0 = non-irrigated (control). I1 = irrigation at vegetative stage. I2 = irrigation at heading stage I3 = irrigation at flowering stage I4 = I1 + I3 (two irrigations). I5 = I1 + I2 + I3 (three irrigations). I6 = I1 + I2 + I3 + irrigation at milking stage were applied. |
I5 and I6 are characterized by the lowest Oil content. I0 marked the highest oil content. I3 the most preferred water regime. |
|
Jordan et al. 2012 |
Two varieties of sunflower (Helio 358 and Aguará 04). |
-Four treatments L1, L2, L3, L4 with different water depths. -Control (rainwater 448mm-454mm). |
-Irrigation increases led to increases in oil content. -Oil content gains in response to irrigation were more pronounced for Aguará 04 variety. |
|
Salem and Ali 2012 |
7 inbred Lines (L38 and L11from Egypt and L350, L460, L990, L770 and L235 from Bulgaria). |
Three levels of water regime: -control supplemented (3000 m³) -moderate drought (2000 m²) -severe drought (1000 m³). |
-L38, L990 and L235 selected for its general combining ability (GCA) for oil content in severe drought. - L38 and L11 and L235 are good combiners for drought tolerance. -The cross combination L38 x L350 was found to promising for seed yield and oil content. |
|
Salem et al.2013 |
Twelve sunflower genotypes. |
Three regimes: -adequate (7140 m3/ha) -moderate (4760 m3/ha) -severe (2380 m3/ha). |
-Moderate and severe levels of drought affect oil content of all the sunflower genotypes differently. -adequate water level increases the oil content in L990 and Giza102. -The oil content of L38 is the best under moderate and severe drought conditions. -L990 is recommended for adequate water supplies and L38 is recommended for moderate and severe water regimes. |
crop evapotranspiration (ETc), field capacity (FC), oleic genotype (Go), standard genotype (Gs), linoleic acid (LA), oleic acid (OA), oleic acid/linoleic acid ratio (OA/LA), palmitic acid (PA), stearic acid (SA)
The fatty acids were analysed by gas chromatography. The transesterification of triglycerides to fatty acid methyl esters was performed with methanoleic potassium hydroxide using a method described by the A.O.A.C. (2002). A capillary column (HP-5) and 50-cm long 5 % methyl phenyl siloxane on an Agilent gas chromatograph (Agilent Technologie 2890 N, Net work GC System) with a flame ionization detector (FID) were used. Injector and detector temperatures were kept at 270 °C and 320 °C, respectively. The carrier gas was helium at a flow rate of 1 ml/min. An electronic integrator (GC-Chemstation, Rev.A.09.01 [1206]) was used to calculate the total area of the peaks. The area of each fatty acid peak was expressed as a percentage of the total area using the following formula:
% fatty acid = Air of fatty acids × 100 / Total air of fatty acids
The experiment was conducted in triplicate. Analysis of variance (ANOVA) was conducted using PROC ANOVA of SAS (version 9.0) and significant differences between means were assessed by the Fisher test at P < 0.01 and 0.05 (Fisher, 1954). Analysis of the interaction between genotype and watering regime and its impact on oil content was performed with the macro PDMix 800 of PROC Mixed for SAS (version 9.0).
3. Results and discussion
Although several studies have focused on sunflower varieties from European Mediterranean countries, this is the first study that has focused on genotypes maintained and cultivated by Tunisian farmers. To improve the sunflower crop in Tunisia, the present research has clearly shown the ability of six lines to resist water shortage during critical stages. By focusing on qualitative traits, the effect of water supply and genotype on oil content and fatty acid composition were analyzed.
3. Effect of watering regime and line on seed oil content
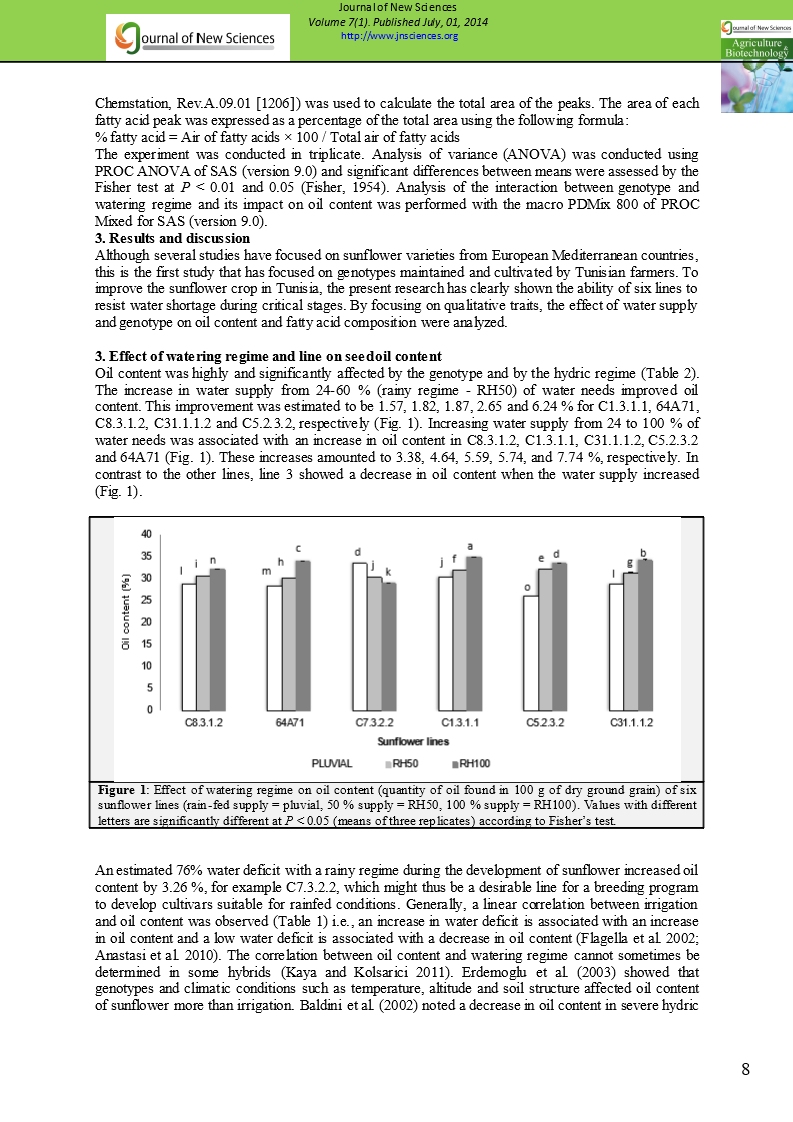
Oil content was highly and significantly affected by the genotype and by the hydric regime (Table 2). The increase in water supply from 24-60 % (rainy regime - RH50) of water needs improved oil content. This improvement was estimated to be 1.57, 1.82, 1.87, 2.65 and 6.24 % for C1.3.1.1, 64A71, C8.3.1.2, C31.1.1.2 and C5.2.3.2, respectively (Fig. 1). Increasing water supply from 24 to 100 % of water needs was associated with an increase in oil content in C8.3.1.2, C1.3.1.1, C31.1.1.2, C5.2.3.2 and 64A71 (Fig. 1). These increases amounted to 3.38, 4.64, 5.59, 5.74, and 7.74 %, respectively. In contrast to the other lines, line 3 showed a decrease in oil content when the water supply increased (Fig. 1).
|
|
|
Figure 1: Effect of watering regime on oil content (quantity of oil found in 100 g of dry ground grain) of six sunflower lines (rain-fed supply = pluvial, 50 % supply = RH50, 100 % supply = RH100). Values with different letters are significantly different at P < 0.05 (means of three replicates) according to Fisher’s test. |

An estimated 76% water deficit with a rainy regime during the development of sunflower increased oil content by 3.26 %, for example C7.3.2.2, which might thus be a desirable line for a breeding program to develop cultivars suitable for rainfed conditions. Generally, a linear correlation between irrigation and oil content was observed (Table 1) i.e., an increase in water deficit is associated with an increase in oil content and a low water deficit is associated with a decrease in oil content (Flagella et al. 2002; Anastasi et al. 2010). The correlation between oil content and watering regime cannot sometimes be determined in some hybrids (Kaya and Kolsarici 2011). Erdemoglu et al. (2003) showed that genotypes and climatic conditions such as temperature, altitude and soil structure affected oil content of sunflower more than irrigation. Baldini et al. (2002) noted a decrease in oil content in severe hydric stress, while a moderate water deficit increased oil content, showing the great adaptability of sunflower to early hydric stress applied during flowering to seed filling. The authors attributed this result to abscisic acid produced in leaves of stressed plants and then translocated to the seed, thus contributing to the decline in seed oil content. Santonoceto et al. (2003) similarly noted that an increase in oil content in standard genotypes was positively correlated with the amount of irrigation supplied (33-100 % of water needs of the crop).
The effect of water supply (Table 1) on seed oil content is due to different experimental conditions such as intensity, timing and level of water stress (Roche et al. 2006). Oil content also differs from line to line and this variation is most likely due to a genotypic effect (Mrinkovic et al. 2011). Tang et al. (2006) fortified this notion by identifying microsatellites markers (“ORS407” and “ORS337”) on linkage groups 16 and 4, respectively, which are associated with the quantitative trait loci (QTLs) related to oil content. These regions are important and may control oil content in water stressed and well watered conditions.
3.2. Effect of watering regime and line on composition of saturated and unsaturated fatty acids
Linoleic and stearic acids were affected by water regime (Table 2). The increase in water supply was associated with a decrease in linoleic acid content, and in comparison to the rainy regime, the decrease varied from 9.34 to 13.52 %, respectively in RH50 and RH100 (Fig. 2). While linoleic content increased as water supply became reduced, the oleic acid content was not significantly affected by regime or genotype (Table 2).

|
Table 2: Mean square of variance analysis, effect of lines and watering regime on fatty acids and oil content (%). |
||||||
|
SV |
DF |
%H |
%P |
%L |
%O |
%S |
|
REG |
2 |
38.31** |
6.23 |
694.08* |
84.73 |
9.21* |
|
Genotype |
5 |
15.65** |
14.91* |
224.34 |
45.20 |
0.98 |
|
REG×Genotype |
10 |
20.80** |
7.36 |
193.19 |
60.79 |
2.38 |
|
Error |
30 |
0.002 |
4.45 |
154.88 |
41.72 |
1.87 |
Stearic acid content (Fig. 3) was affected by water stress (Table 2). In comparison to RH50, more stearic acid was obtained in the rainy regime but less than RH100. The rainy regime showed continuous stress during the different phases of the development of sunflowers (36.95 % of water needs during the vegetative – flowering phase and 18.66 % of water needs during late flowering – maturity phase was provided). This rainy regime induced an increase in stearic acid content of 0.22 % compared to the RH50 regime, providing 60 % of the water needs of sunflower.
|
|
|
Figure 3: Effect of watering regime (rain-fed supply = pluvial, 50 % supply = RH50, 100 % supply = RH100) on stearic acid content (average of six lines). Values with different letters are significantly different at P < 0.05 (means of three replicates) according to Fisher’s test; error bars = SD. |

Palmitic acid content was significantly affected by genotype. Line C8.3.1.2 had the highest palmitic acid content (6.04 %) while line C7.3.2.2 had the lowest content (4.19 %) (Fig.4). Palmitic acid content for the remaining four lines ranged between 4.40 and 6.04 %. Unlike phasic stress, stress applied when there was rainfall during the different phases of sunflower development (36.95 % of water needs during the vegetative – flowering phase and 18.66 % of water requirements during flowering – maturity), induced an increase in C18:1/C18:2 ranging from 0.20 to 0.60. The RH50 regime provided 60 % of water needs during the final phase of development (flowering – maturity) and was also marked by an increase in C18:1/C18:2 ranging between 0.30 and 0.50. The fatty acid composition is known to differ between cultivars under different environmental conditions (Baldini et al. 2000). Water supply, genotype and temperature during oil formation exert a major effect on the proportions of oleic and linoleic acids (Roche, 2005). Different fatty acids (18:0, 18:1, 16:0 and 18:2) indicate variations between genotypes when plants are exposed to different water regimes, namely irrigated and non-irrigated (Flagella et al. 2002).
|
|
|
Figure 4: Palmitic acid content of six sunflower lines. Values with different letters are significantly different at P < 0.05 (means of three replicates) according to Fisher’s test; error bars = SD; line 1 = C8.3.1.2; line 2 = 64A71; line 3 = C7.3.2.2; line 4 = C1.3.1.1; line 5 = C5.2.3.2; line 6 = C31.1.1.2 |

As noted in different studies (Table 1) conducted on standard and oleic genotypes (Piva et al. 2000; Baldini et al. 2002; Anastasi et al. 2010; Sezen et al. 2011), linoleic (C18:2) and oleic (C18:1) acids respond differently to water deficit, and usually an increase in water deficit is associated with an increase in C18:1 and a decrease in C18:2 therefore an increase in the C18:1/C18:2 ratio. This ratio is affected by the timing of stress application such that a ratio of 2.09 was observed when hydric stress was applied at an early stage during flowering to seed filling while a 1.52 ratio was observed when the stress was applied late during seed filling to seed maturity (Baldini et al. 2002). Under severe stress (33 % and 67 % ETc), Anastasi at al. (2010) noted an 8.9 % reduction in oleic acid and a 8.5 % increase in linoleic acid in the oil of standard and oleic genotypes. Oleic acid in high oleic hybrids was among the main fatty acids that increased (2.8 %) under severe hydric stress in which water supply was not applied at the most critical stages of sunflower seed yield (flower bud appearance and flowering), accentuating the influence of genotype (Flagella et al. 2002). Salera and Baldini (1998) showed no effect of water management on oleic acid content both in standard and high oleic genotypes, with oleic acid content not being significantly affected by the watering regime or genotype.
These variations might be caused by enzymatic processes governed by genes coding for oleoyl ∆-9 desaturase and oleoyl ∆-12 desaturase, which are in fact being the main enzymes implicated in metabolic processes related to oleic and linoleic acids, respectively (Cantisan et al. 2000; Rondanini et al. 2003; Lacombe et al. 2004). Thus, variation in the proportion of unsaturated fatty acid (C18:2) could be explained by a different strategy under drought conditions that implicates an up- or down-regulation of ∆-12 desaturase (Anastasi at al. 2010).
Baldini et al. (2002) also noted that water stress usually accelerates earlier embryo development and stimulates enzymatic activities of fatty acid biosynthesis, including oleoyl ∆-12 desaturase which could alter the final acid composition of sunflower seeds.
Roche et al. (2006) and Baldini et al. (2002) reported that variation in oleic acid may be due to the synthesis or activation of oleate desaturase, which is affected by the irrigation supply. The cellular structure and probably nutritional status under non-irrigated conditions may affect the activation or synthesis of oleate desaturase. The synthesis of 18:1 in seeds occurs in proplastids and synthesis of 18:2 and 18:3 from 18:1 occur in the cytosol (Dossat et al. 2002). Thus, environmental factors may influence the proportions of fatty acids not only by altering enzyme activity but also by influencing the transport of fatty acid from one organelle to another (Roche, 2005).
According to several authors (Lacombe and Berville 2001; Martinez-Rivas et al. 2001; Pérez-Vich et al. 2002b; Schuppert et al. 2006), variation in unsaturated fatty acids is governed by different genes in standard and oleic genotypes that responds to different environment conditions. Flagella et al. (2002) and Sezen et al. (2011) also noted that stearic content was significantly affected by soil water content. That is, water stress during the seed-filling stage increased the percentage of stearic acid but an irrigation-based regime significantly decreased the stearic acid content.
According to Flagella et al. (2002), palmitic acid content is mainly affected by water regime. This is confirmed by the results of our study: under irrigation, in which water was applied at the most critical stages of seed yield (flower bud appearance and flowering), palmitic acid increased and in the non-irrigation regime (i.e., based only on rainfall and under water deficit), palmitic content decreased (Table 1).
4. Conclusion
Among the different genotypes and stress conditions imposed in our experimental design, line 3 is characterized by the lowest palmitic acid content and a decrease in oil content when water supply increases. Thus, line 3 may be a desirable line for a breeding program to develop cultivars suitable for rain-fed conditions. While changes in palmitic, stearic and linoleic acids were similar to trends observed in other studies in the literature, in this study, the oleic acid content marked a key difference by not being significantly affected by the line or by the hydric regime. Despite this, further studies are required to understand the mechanisms implicated in the oleic acid content in response to different water regimes. Genotype remains the most important factor controlling the variability in oil and fatty acid composition, mainly palmitic acid. Water supply had a prominent effect on oil percentage, as well as saturated and unsaturated fatty acid composition, thus the choice of genotype and conditions would be useful in designing management practices to obtain a specific oil quality.
5. Acknowledgments
The authors are grateful to INAT (National Institute of Agronomy of Tunisia) for funding this study and to INRAP (Laboratory of Methods and Analytical Techniques, National Institute for Research and Physico-chemical Analysis) for GC analyses of the oil samples.
6. References
Anastasi U, Cammarata M, Abbate V (2000) Yield potential and oil quality of sunflower (oleic and standard) grown between autumn and summer. Italian Journal of Agronomy 4: 23–36.
Anastasi U, Santonoceto C, Giuffrè, A.M, Sortino O, Gresta F, Abbate V (2010) Yield performance and grain lipid composition of standard and oleic sunflower as affected by water supply. Field Crops Research 119: 145–153.
Association Française de Normalisation (AFNOR) (1998) NF EN ISO 659. Norme 255 francaise V03-905 Graine oléagineuses: Détermination de la teneur en huile (méthode de référence) [Seed oil: Determination of oil content (Reference method)]. Paris
Association of Official Analytical Chemists (AOAC) (2002) Official methods of analysis of AOAC International, 17 th ed. Gaithersburg, MD: AOAC.
Baldini M, Giovanardi R, Tahmasebi-enferadi S, Vannozzi G (2002) Effects of water regime on fatty acid accumulation and final fatty acid composition in the oil of standard and high oleic sunflower hybrids. Italian Journal of Agronomy 6: 119–126.
Boyeldieu J. (1991) Le tournesol. In: Moati. P. (ed.), Produire des grains protéagineux et oléagineux. Lavoisier-Tec et Doc, Condé-sur-Noireau: 68-113.
Cantisan S, Martínez-Force E, Garcés R (2000). Enzymatic studies of high stearic acid sunflower seed mutants. Plant Physiology and Biochemistry 38: 377-382.
Centre Technique Interprofessionnel des Oléagineux Métropolitains (CETIOM) and La Filière Française des Huiles Et Protéines Végétales (PROLEA) (2008) Irrigation. 285 http//www.CETIOM et PROLEA.fr.
CODEX Stan 210 (1999) Codex Standard for named vegetable oils. Current official standards (Amended 2003, 2005). FAO/WHO Food standard. Codex Alimentarius. http://www.codes alimentarius net. Last accessed: 25 June, 2014
Dossat V, Combes D, Marty A (2002) Lipase-catalysed transesterification of high oleic sunflower oil. Enzyme and Microbial Technology 30: 90–94.
Echarte M.M, Angeloni P, Jaimes F, Tognetti J, Izquierdo NG, Valentinuz OAguirrezabal LAN (2010) Night temperature and intercepted solar radiation additively contribute to oleic acid percentage in sunflower oil. Field Crops Research 119: 27–35.
Erdemoglu N, Kusmenoglu S, Yenice N (2003) Effect of irrigation on the oil content and fatty acid composition of some sunflower seeds. Chemistry of Natural Compounds 39: 1-4.
Fisher RA (1954) Statistical methods for research workers. Oliver and Boyd. ISBN 0-05-002170-2.
Flagella Z, Giuliani M.M, Rotunno T, Di Caterina R, De Caro A (2004) Effect of saline water on oil yield and quality of a high oleic sunflower (Helianthus annuus L.) hybrid. European Journal of Agronomy 21: 267–272.
Flagella Z, Rotunno T, Di Caterina R, De Simone G, De Caro A (2000) Effect of environmental crop conditions to produce useful sunflower oil components. Meteorology 143: 252–265.
Flagella Z, Rotunno T, Tarantino E, Di Caterina R, De Caro A (2002) Changes in the seed yield and fatty acid composition of high oleic sunflower (Helianthus annuus L.) hybrids in relation to sowing date and water regime. European Journal of Agronomy 17: 221–230.
Garcés R, Alvarez-Ortega R, Cantisan S, Martínez-Force E (2000) Biochemical control of high palmitic acid biosynthesis. In: Proc. XV International Sunflower Conference, Toulouse, France, IA: 7-12.
Izquierdo N, Aguirrezabal L, Andrade F, Pereyra V (2002) Night temperature affects fatty acid composition in sunflower oil depending on the hybrid and the phonological stage. Field Crops Research 77: 115–126.
Izquierdo N.G, Aguirrezábal L.A.N (2008) Genetic variability in the response of fatty acid composition to minimum night temperature during grain filling in sunflower (Helianthus annuus L.) exposed to brief intervals of high temperature. European Journal of Lipid Science and Technology 108: 287–297.
Kaya MD, Kolsarici O (2011) Seed yield and oil content of some sunflower (Helianthus annuus L.) hybrids irrigated at different growth stages. African Journal of Biotechnology 10: 4591–4595.
Kazemeini SA, Edalat M, Shekoofa A (2009) Interaction effects of deficit irrigation and row spacing on sunflower (Helianthus annuus L.) growth, seed yield and oil yield. African Journal of Agricultural Research 11: 1165–1170.
Lacombe S, Bervillé A (2001) A dominant mutation for high oleic acid content in sunflower (Helianthus annuus L.) seed oil is genetically linked to a single oleate desaturase RFLP locus. Molecular Breeding 8: 129–137.
Lacombe S, Kaan F, Griveau A, Bervillé A (2004) The prevents high oleic mutation: methodological studies. Helia 27: 41–54.
Marinković R, Jocković M, Marjanović-Jeromela A, Jocić S, Ćirić M, Balalić I, Sakač Z (2011) Genotype by environment interactions for seed yield and oil content in sunflower (H annuus L.) using AMMI model. Helia 34: 79–88.
Martinez-Rivas JM, Sperling P, Lühs W, Heinz E (2001) Spatial and temporal regulation of three different microsomal oleate desaturase genes (FAD2) from normal type and high-oleic varieties of sunflower (Helianthus annuus L.). Molecular Breeding 8: 159–168.
Pereira-Irujo GA, Aguirrezábal LAN (2007) Sunflower yield and oil quality interactions and variability: analysis through a simple simulation model. Agricultural and Forest Meteorology 143: 252–265.
Pérez-Vich B, Garcés R, Fernández-Martínez JM (2002a) Inheritance of high palmitic acid and its relationship with high oleic acid content in the sunflower mutant CAS-12. Plant Breeding 121: 49–56.
Pérez-Vich B, Garcés R, Fernández-Martínez JM (2002b) Inheritance of medium stearic acid content in the seed oil of sunflower mutant CAS-4. Crop Science 42: 1806–1811.
Pérez-Vich B, Leon AJ, Grondona M, Velasco L, Fernández-Martínez JM (2006) Molecular analysis of the high stearic acid content in sunflower mutant CAS-14. Theoretical and Applied Genetics 112: 867–875.
Pérez-Vich B, Velasco L, Fernández-Martínez JM (1998) Determination of seed oil content and fatty acid composition in sunflower through the analysis of intact seeds, husked seeds, meal and oil by near-infrared reflectance spectroscopy. Journal of the American Oil Chemists’ Society 75: 547–555.
Petcu E, Arsintesscu A, Stanciu D (2001) The effect of drought stress on fatty acids composition in some Romanian sunflower hybrids. Romanian Agricultural Research 15: 39–43.
Piva G, Bouniols A, Mondiès M (2000) Effect of cultural conditions on yield, oil content and fatty acid composition of sunflower kernel. In: XV International Sunflower Conference, Toulouse, France, IA: 61-66.
Roche J (2005) Composition de la graine de tournesol (Helianthus annuus L.) sous l’effet conjugué des contraintes agri-environnementales et des potentiels variétaux. PhD thesis, Institut National Polytechnique de Toulouse, France: 305.
Roche J, Bouniols A, Moulounguib Z, Barrancoa T, Cernyb M (2006) Management of environmental crop conditions to produce useful sunflower oil components. European Journal of Lipid Science and Technology 108: 287–297.
Rondanini D, Mantese A, Savin R, Hall AJ (2006) Responses of sunflower yield and grain quality to alternating day/night high temperature regimes during grain filling: Effects of timing, duration and intensity of exposure to stress. Field Crops Research 96: 48–62.
Rondanini D, Savin R, Hall AJ (2003) Dynamics of fruit growth and oil quality of sunflower. Field Crops Research 106: 116–125.
Salera E, Baldini M (1998) Performance of high and low oleic acid hybrids of sunflower under different environmental conditions, Note II. Helia 21: 55–68.
Santonoceto C, Anastasi U, Riggi E, Abbate V (2003) Accumulation dynamics of dry matter, oil and major fatty acids in sunflower seeds in relation to genotype and water regime. Italian Journal of Agronomy 7: 3–14.
SAS Institute (2002) The SAS Systemfor Windows. Release 9.0. SAS Inst Inc., Cary, NC, USA.
Schuppert GF, Tang S, Slabaugh MB, Knapp SJ (2006) The sunflower high oleic mutant Ol carries variable tandem repeats of FAD2-1, a seed specific oleoylphosphatidyl choline desaturase. Molecular Breeding 17: 214–256.
Sezen SM, Yazarb A, Kapurb B, Tekin S (2011) Comparison of drip and sprinkler irrigation strategies on sunflower seed and oil yield and quality under Mediterranean climatic conditions. Agricultural Water Management 98: 1153–1161.
Tang S, Leon A, Bridges WC, Knapp SJ (2006) Quantitative trait loci for genetically correlated seed traits are tightly linked to branching and pericarp pigment loci in sunflower. Crop Science 46: 721–734.
Velasco L, Pérez-Vich B, Fernández-Martínez JM (2000) Inheritance of oleic acid contents under controlled environment. In: Proc. 15th international sunflower conference, 12-15, 1: PL. AS-10.
